Abstract
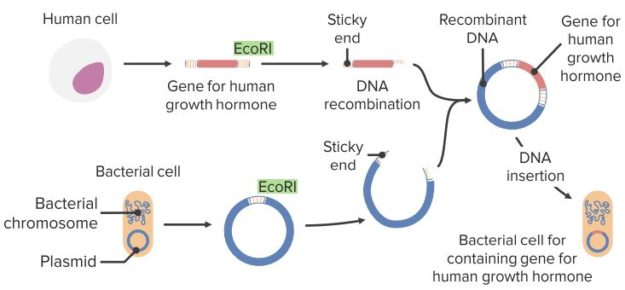
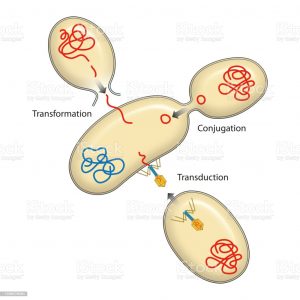
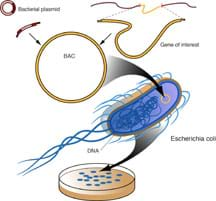
The discoveries of the double-stranded structure of DNA and the development of recombinant DNA technology sparked a revolution in industrial microbiology. Traditional industrial microbiology merged with molecular biology to produce improved organism recombinant processes for the industrial production of primary and secondary metabolites, biopharmaceutical proteins, and industrial enzymes. New genetic techniques such as metabolic engineering, combinatorial biosynthesis and molecular breeding techniques and their modifications are greatly contributing to the development of improved industrial processes.
Furthermore, functional genomics, proteomics and metabolomics are being exploited for the discovery of new small molecules valuable for medicine, as well as enzymes for catalysis. Sequencing of industrial microbial genomes is ongoing, which bodes well for future process improvement and the discovery of new industrial products.
Keywords:
antibiotics, antitumor agents, biopharmaceuticals, hypocholesterolemic agents, enzymes, genetic recombination, industrial products, primary metabolites, secondary metabolites
Amino acids.
Genetic engineering has impacted amino acid production through the use of the following strategies: (1) amplification of a rate-limiting enzyme pathway; (2) amplification of the first enzyme after a branch point; (3) cloning of a gene encoding an enzyme with more or less feedback regulation; (4) introduction of a gene encoding an enzyme with a functional or energetic advantage as a replacement for the normal enzyme; and (5) amplification of the first driving enzyme from central metabolism to increase carbon flux in the pathway followed by sequential removal of bottlenecks caused by the accumulation of intermediates.
Transport mutations have also become useful, that is, a mutation that reduces amino acid uptake allows for enhanced excretion and lower intracellular feedback control. This has been especially useful in the production of tryptophan and threonine. In cases where excretion is mediated by transporters, increased activity of these transporter enzymes increases amino acid production.
Other compounds.
Nucleosides. Genetic engineering of the inosine monophosphate (IMP) dehydrogenase gene in a B. subtilis strain that produces 7 gl−1 desirable guanosine and 19 gl−1 undesirable inosines changed the production to 20 gl−1 guanosine and 5 gl −1 inosine. Guanosine is used to produce guanosine monophosphate (GMP), a powerful flavouring agent.
Carotenoids. Carotenoids were overproduced by the introduction of Erwinia uredovora carotenoid gene clusters into E. coli and overexpression of E. coli deoxyglucose phosphate synthase, the key enzyme of the non-mevalonate isoprenoid biosynthetic pathway zeaxanthin at 0.6 mg g−1.
Glucosamine. Glucosamine is used for osteoarthritis and is produced by acid hydrolysis of chitin from shellfish waste. Since many patients have shellfish allergies, a microbial source is desirable. Metabolic engineering of E. coli produced a process that produced 17 gl−1 of glucosamine. In this strain, glucosamine synthase was overexpressed, glucosamine-degrading genes were inactivated, and glucosamine inhibition of glucosamine synthase was decreased. by mutational modification of the enzyme through error-prone PCR. Overexpression of a heterologous glucosamine-6-P-N acetyltransferase produced a strain that produces 110 g l -1 of N-acetylglucosamine which is readily converted to glucosamine by mild acid hydrolysis.
Solvents. Cloning it’s as (acetone) ADC operon genes (encoding acetoacetate decarboxylase), and CTF and CTF (two genes encoding coenzyme A transferase) into a plasmid containing the ADC promoter in Clostridium acetobutylicum resulted in a 95% increase % in the production. The increases were 95% in acetone production, 37% in butanol, 90% in ethanol, 50% yield of solvent from glucose, and 22-fold lower production of undesirable acids. The introduction of the acetone operon of C acetobutylicum in E. coli led to the high production of acetone by the latter.